Зрительный аппарат и цветовое зрение
Сигналы о величи нах каждой из них передаются в мозг V, вызывая соответственно ощущения красного, зеленого и синего цветов. Эти элементарные ощущения называются основными. Они складываются в единое, зависящее от соотношения основных. Например, при преобладании реакций рецепторов, ответственных за возникновение «зеленых» сигналов, появляется ощущение зеленого цвета. Если перевес на стороне «зеленых» и «красных» одновременно, возникает ощущение желтого. Соотношение интенсивностей основных сигналов определяется спектральным составом излучения и формой характеристик 4, 5 и 6. Если бы они были узкими и не перекрывались, то для описания цвета достаточно было бы знать спектральный состав излучения. Вследствие взаимного перекрывания кривых излучения разных спектральных составов могут давать одинаковые соотношения интенсивностей сигналов и, следовательно, вызывать ощущения одинаковых цветов.
Процесс формирования цветового ощущения осложняется психологическими факторами, которые будут рассмотрены в главе 3.
2.1. Общие сведения о зрительном аппарате
Орган зрения в целом состоит из трех отделов — периферического (собственно глаз), проводникового (зрительный нерв) и центрального (зрительная зона коры головного мозга в затылочной области).
Рассмотрим в общих чертах строение глаза, опуская детали, имеющие для теории цвета второстепенное значение.
Глазная линза — хрусталик — дает оптическое изображение наблюдаемого предмета, которое системой нервных окончаний, находящихся в одной из оболочек глаза, преобразуется в сигналы. Они по зрительному нерву передаются в затылочные доли головного мозга. В результате этого по неизвестным пока механизмам возникает зрительный образ предмета.
На рис. 2.1 схематически показан разрез глаза. Он представляет собой шарообразное тело, образованное несколькими оболочками. Внешняя /, называемая белковой оболочкой или склерой, состоит из сухожилий, непрозрачна и выполняет защитную роль.
Спереди она переходит в прозрачную и более выпуклую оболочку 2 — роговую. Под склерой находится сосудистая оболочка 3, в которой заключены кровеносные сосуды, питающие глаз. К ней по внутренней сто роне примыкает пигментный слой клеток (на рисунке совпадает с внутренним контуром разреза сосудистой оболочки). Клетки поглощают рассеянный свет. Пигментный слой предохраняет оптическое изображение, создаваемое глазной линзой — хрусталиком 6, от чрезмерного искажения рассеянным светом. Сосудистая оболочка спереди переходит в ресничное (цилиарное) тело 4, а затем — в радужную оболочку 5, или радужку, содержащую пигментные клетки. Пространство между хрусталиком 6 и роговой оболочкой (передняя камера 7) заполнено так называемой водя»истой влагой. Она преимущественно состоит из воды (99%), в которой растворены соли и белки. За хрусталиком находится стекловидное тело 8, также состоящее главным образом из воды.
Отверстие в центре радужки — зрачок — играет роль диафрагмы. При изменении светового потока, попадающего в глаз, площадь зрачка меняется: либо круговые мышцы радужки сужают его, либо радиальные расширяют. Эти реакции (зрачковый рефлекс) непроизвольны, и их роль заключается в предохранении светочувствительной оболочки глаза — сетчатки от чрезмерного раздражения при повышении освещенности. При ее снижении зрачковый рефлекс обеспечивает достаточную чувствительность оболочки.
Если в оптических приборах наводка на резкость осуществляется изменением расстояния от объектива до оптического изображения, то в органе зрения аналогичный процесс происходит путем изменения оптической силы хрусталика, определяемой кривизной его поверхностей. Кривизной управляют мышцы ресничного тела 4, находящегося в основании радужной оболочки. При сокращении круговых мышц уменьшается натяжение связок 9 хрусталика, называемых цинновыми. Тогда упругий хрусталик принимает естественную для него выпуклую форму, фокусное расстояние уменьшается и близкий предмет изображается резко.
Если же предмет удален, круговые мышцы ресничного тела расслабляются, а радиальные сокращаются. В результате этого хрусталик становится менее выпуклым и его фокусное расстояние возрастает. Эти явления получили название аккомодации.
Сетчаткой (ретиной, или сетчатой оболочкой) называется внутренняя оболочка 10. Это — светочувствительный слой глаза. В сетчатке находятся нервные окончания (рецепторы) в которых происходят начальные преобразования лучистой энергии, приводящие в конце концов к возникновению светового ощущения.
Из глаза выходит зрительный нерв 11, по которому нервные импульсы, возникающие вследствие обратимого фотораспада веществ, находящихся в рецепторах, передаются в мозг. Место выхода зрительного нерва — слепое пятно 12—участок, не содержащий рецепторов.

Рис. 2.1. Схема строения глаза:
1 — склера; 2 — роговица; 3 — сосудистая оболочка; 4 — ресничное тело; 5 — радужная оболочка; 6 — хрусталик; 7 — передняя камера; 8 — стекловидное тело; 9 — цинно-вы связки; 10 — сетчатка; 11 — зрительный нерв; 12 — слепое пятно; 13 — желтое пятно; 14 — центральная ямка; I5 — зрительная ось; 16 — оптическая ось

Рис. 2.2. Схема строения сетчатки:
1, 2, 3 — слои нейронов; а — нервные волокна; б — синапсы; в — палочки; г — колбочки; д — пигментный эпителий; е — биполярные клетки; ж — ганглии
В сетчатке — три слоя нервных клеток (рис. 2.2) — нейронов /, 2, 3, связанных разветвлениями—синапсами б, обеспечивающими передачу электрического сигнала от одной клетки к другой. Нейроны, наиболее удаленные от внутренней поверхности сетчатки, оканчиваются рецепторами в и г. Они бывают двух типов: длинные и тонкие называются палочками (в), толстые и короткие — колбочками (г). Палочки обеспечивают черно-белое зрение, колбочки -как черно-белое, так и цветное. Шестиугольные по форме пигментные клетки (рис. 2.2) охватывают своими отростками рецепторы (на рисунке не показано).
Рецепторы передают сигнал через биполярные клетки е второго слоя ганглиям ж (скопления нервных волокон), от которых он попадает в зрительный нерв.
Наиболее важная с точки зрения цветовосприятия область сетчатки -- желтое пятно 13 (рис. 2.1), расположенное в центральной ее части. Оно окрашено желтым пигментом, предохраняющим рецепторы этой области от чрезмерного возбуждения коротковолновыми излучениями. Средняя часть 14 желтого пятна углублена и называется поэтому центральной ямкой. В середине центральной ямки находится область, содержащая только колбочки. Она имеет угловой размер около 2°, что соответствует площади меньше 1 мм2. Здесь насчитывается около 50 тыс. колбочек, очень близко расположенных друг к другу. Высокая поверхностная концентрация рецепторов обеспечивает большую разрешающую способность и цветовую чувствительность этого участка сетчатки.
При наблюдении детали предмета глаз ориентируется так, чтобы ее изображение упало на середину ямки. Такая ориентация обеспечивает наилучшее восприятие. Прямая, соединяющая центр ямки с наблюдаемой точкой предмета, как говорят, точкой фиксации взора, называется зрительной осью 15. При рассмотрении предмета в целом глаз движется. Он принимает разные положения, и оптические изображения деталей объекта, привлекающих внимание наблюдателя, поочередно проецируются на центральный участок ямки. Глаз «ощупывает» им наблюдаемый предмет. Вследствие подвижности глаза, наблюдатель не испытывает неудобств от того, что наиболее полезный участок сетчатки очень мал. В тех случаях, когда при исследованиях или измерениях хотят, чтобы работал только центральный участок ямки, угол зрения ограничивают соответствующим образом.
С удалением от средней части центральной ямки растет концентрация палочек и падает количество колбочек, приходящихся на единицу площади сетчатки. Изображение, образующееся на периферической ее части, не дает подробной информации об объекте. Оно позволяет лишь ориентироваться в пространстве.
Световая чувствительность палочек и колоочек резки различна. Палочки работают при низких освещенностях и выключаются при высоких.
Эти рецепторы обеспечивают так называемое сумеречное зрение, когда освещенности невелики. В полутьме не различаются цвета, плохо видны детали. Это объясняется тем, что палочки располагаются на сетчатке значительно реже, чем колбочки, и разрешающая способность палочкового аппарата намного ниже - чем колбочкового. Однако в сумерках человек может ориентироваться, получая общее представление о предметах внешнего мира.
Колбочковое зрение называется дневным. При высоких освещенностях, когда начинают действовать колбочки, глаз различает цвета и мелкие детали объектов.
При некоторых средних освещенностях (так называемых промежуточных), когда яркости окружающих предметов находятся в пределах 0,01 — 10кд·м-2, палочки и колбочки работают совместно.
В результате светового возбуждения палочек или колбочек в мозг передаются электрические импульсы, частота которых увеличивается с ростом освещенности сетчатки. Импульсы достигают затылочных долей мозга, где возбуждают световые ощущения, из которых складывается зрительный образ объекта.
Причина возникновения импульсов состоит в фотодиссоциации светочувствительных пигментов, заключенных в рецепторах. Палочки содержат пигмент, называемый родопсином (или зрительным пурпуром), который в результате освещения обратимо распадается. Продуктами распада служат ионы белков — протеина и ретинена. По мере распада пигмента в палочках накапливается отрицательный заряд, и когда он достигает порогового значения, в нервном волокне возникает импульс, передаваемый в зрительную зону коры мозга. Частота импульсов растет с возрастанием освещенности сетчатки. После прекращения импульса происходит регенерация родопсина. Процесс этот идет при участии пигментного слоя д (рис. 2.2), в контакте с которым находятся палочки. Источником энергии, необходимой для протекания реакций, служит кислород, доставляемый кровью к тканям глаза.
Процессы, протекающие в колбочках, менее изучены, хотя несомненно, что их сущность также состоит в фотодиссоциации зрительного пигмента и возникновении электрических импульсов.
Со времени М. В. Ломоносова предполагалось, а в середине 20 века нашло экспериментальное подтверждение, что колбочки неодинаковы по спектральным свойствам, это позволяет разделить их на три группы. При возбуждении рецепторов первой группы возникает ощущение сине-фиолетового цвета. Рецепторы этой группы называются синечувствительными. Рецепторы второй группы ответственны за ощущение зеленого. Третья группа рецепторов обеспечивает ощущение красного. Реальные излучения раздражают колбочки сразу нескольких групп. Комбинация раздражений дает ощущения голубого, оранжевого и других цветов.
Р. Гранит экспериментально показал, что сетчатка животных дает три рода реакций. Экспериментатор удалял переднюю часть глаза кошки и тонкими (0,25 мкм) электродами касался разных участков обнаженной сетчатки глаза наркотизированного животного. Освещение сетчатки приводило к возникновению на электродах импульсов тока. Оказалось, что их величина и частота зависят от длины волны излучения и яркости света. В результате опытов были получены три кривые спектральной чувствительности, подтвердившие существование колбочек трех типов.
У. Раштону удалось доказать существование двух светочувствительных пигментов в колбочках. Возбуждающий ощущение зеленого был им назван хлоролабом («захватывающий зеленый»), красного — эритролабом («захватывающий красный»). По мнению исследователя, существует и третий пигмент — цианолаб («захватывающий синий»), световая диссоциация которого приводит к возникновению ощущения синего цвета.
Вопрос о механизме действия разных светочувствитель- ных пигментов не совсем ясен. Существует мнение, что при возбуждении колбочек возникают импульсы, частота которых зависит от спектральной чувствительности соответствующего рецептора. Соотношение частот импульсов определяет цветность излучения.
Важную роль в механизме зрения играют непроизвольные движения глаза. Он не бывает неподвижным даже, когда наблюдателю кажется, что взор фиксирован на определенной точке наблюдаемого предмета.
Схема движений глаза показана на рис. 2.3. На нем представлен участок сетчатки. В центре рисунка — кружок, заключающий оптическое изображение точки, на которой наблюдатель пытается фиксировать взор. Показаны все три типа перемещений глазного яблока при попытке фиксации взора: мелкое дрожание с частотой около 50 Гц и амплитудой, равной приблизительно половине диаметра колбочки,— тремор; перемещение глазного яблока в течение тремора — дрейф; скачкообразные движения (продолжительность, скачка около 20 мс) — саккады.
Если изображение на сетчатке искусственно сделать неподвижным, видимый образ бледнеет и, как правило, частично исчезает. Это объясняется тем. что светочувствительные вещества рецепторов, подвергаемые обратимому фотораспаду, в моменты перемещения восстанавливаются, а в моменты временной остановки движения (углы на рисунке) вновь распадаются под действием света, давая импульсы. Искус-

Рис. 2.3. Схема движений глаза:
I — дрейф; 2 — тремор; 3 — саккада; 4 — желаемое положение фиксации; 5 — граница фактических положений фиксации
ственная фиксация приводит к тому, что зрительные пигменты восстановиться не могут, что ведет к нарушению зрительного процесса.
2.2. ГЛАЗ КАК ВОСПРОИЗВОДЯЩАЯ СИСТЕМА
Глаз — часть живого организма. Механизмы зрения еще во многом неясны. Здесь мы может описать только некоторую модель глаза, иллюстрирующую его основные оптические свойства.
Свойства зрительной системы как воспроизводящей изображение характеризуются ее чувствительностью, постоянством чувствительности, инерционностью.
2.2.1. Световая и спектральная чувствительность глаза
Общие сведения о чувствительности глаза. Под чувствительностью системы или приемника излучения подразумевается их свойство реагировать на излучение. Мера этого свойства, т. е. количественное выражение чувствительности, связана с реакцией приемника на поглощенную световую энергию. Количественно чувствительность выражается как величина, обратная мощности излучения, вызывающей определенную реакцию.
Например, чувствительность фотоэле мента определяют как величину, обратную потоку излучения, вызывающему появление фототока. Чувствительность фотопленки есть величина, обратная экспозиции, дающей определенное почернение.
Следовательно, в общем случае чувствительность глаза S определяется как:
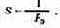
(2.1)
где Рэ — мощность излучения, вызывающая заранее обусловленный зрительный эффект.
Чувствительность, рассчитанная по формуле (2.1), называется абсолютной. В некоторых случаях бывает достаточным выразить чувствительность глаза долей наибольшего значения абсолютной. Эта доля называется относительной чувствительностью. Указанной долей измеряют значения спектральной чувствительности (см. с. 15).
Реакции зрительной системы на излучения могут быть разными. В соответствии с этим различают несколько „типов чувствительности глаза: световую, спектральную, контрастную, чувствительность к цветовому тону, к насыщенности.
Остановимся на световой и спектральной чувствительности, а остальные типы рассмотрим позднее, после введения понятия о порогах цветоразличения.
Световая чувствительность. Способность глаза реагировать на возможно малый поток излучения называется световой чувствительностью. Она измеряется как величина, обратная пороговой яркости. Пороговой называется та наименьшая яркость объекта, например светового пятна, при которой оно может быть обнаружено с достаточной вероятностью на абсолютно черном фоне (т. е. Вфона
= 0). Вероятность обнаружения зависит не только от яркости объекта, но и от угла зрения, под которым он рассматривается, или, как говорят, от его углового размера. С возрастанием углового размера растет число рецепторов, на которые проецируется пятно. Практически, однако, с увеличением угла зрения более чем на 50° чувствительность глаза перестает изменяться.
В соответствии с этим световая чувствительность Sп определяется как величина, обратная пороговой яркости Sп, при условии, что угол зрения а ? ?50°:
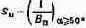
(2.2)
Световая чувствительность очень велика. Так, поданным Н. И. Пинегина, для отдельных наблюдателей минимум энергии, необходимый для появления зрительного эффекта, составляет 3—4 кванта. Это значит, что в благоприятных условиях палочковая световая чувствительность глаза близка к предельной, физически мыслимой.
Колбочковая световая чувствительность, обеспечивающая цветовые ощущения, намного ниже «ахроматической», палочковой. По Н. И. Пинегину, для возбуждения колбоч-кового зрения необходимо, чтобы на одну колбочку в среднем упало не менее 100 квантов.
Спектральная чувствительность. Монохроматические излучения действуют на глаз по-разному. Его реакция максимальна на среднюю часть спектра. Чувствительность к монохроматическим излучениям, определяемая как относительная, называется спектральной.
Реакция глаза, выражающаяся в возникновении светового ощущения, зависит, во-первых, от потока излучения Ф?, упавшего на сетчатку, а во-вторых,— от той доли потока, которая воздействует на рецепторы. Эта доля есть спектральная чувствительность к?. Иногда для обозначения того же понятия применяется термин спектральная эффективность излучения. Произведение к?Ф?, определяет характеристику потока излучения, связанную с уровнем его светового действия и называемую световым потоком F?
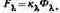
(2.3)
Следовательно, абсолютное значение спектральной чувствительности определяется отношением
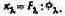
(2.3,а)
Глаз имеет наибольшую спектральную чувствительность к излучению ? = 555 нм, относительно которой определи ются все другие значения этой величины.
Прл световых измерениях значение к?, в формуле (2. 3 принято заменять произведением к555 • ?? где ??
— относительное значение спектральной чувствительности, называемое относительной спектральной световой эффективностью излучения ( в и д н о с т ь ю): ??= к? : к555.
Отсюда абсолютная спектральная чувствительность к?, = к555 • ??
Рис. 2.4. Схема простейшего фотометра
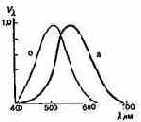
Рис. 2.5. Кривые относительной спектральной световой эффективности:
а — дневное зрение; б — сумеречное зрение
Принцип измерения относительной спектральной чувствительности показан на рис. 2.4. На одну половину фото метрического поля направляют излучение длиной волны 555 нм и известной мощности Ф555, на другую — излучение, спектральная чувствительность к которому измеряется, и имеющее мощность Ф?. С помощью светорегулирующих устройств (на рисунке для простоты — оптический клин) светлоты обеих половин фотометрического поля уравнивают. Это значит, что падающие на них световые потоки F? и F555 становятся равными. Но тогда к?Ф?
= к555 Ф555 и, следовательно,

Однако погрешности результатов сравнения разноокра-шенных половин фотометрического поля велики. Чтобы повысить надежность измерений, используют разные методы. Остановимся на одном из них—способе мелькания (мигания). Он основан на том, что при импульсном освещении
глаз различает изменение светлоты быстрее, чем изменение цветности. Иначе — при очень короткой вспышке ощущается ее интенсивность, но еще не воспринимается цветность, которая станет заметной только при увеличении продолжительности вспышки.
При измерениях по способу мелькания обе половины фотометрического поля освещаются мигающим светом. Частота миганий подбирается так, чтобы еще не различались цветности, но уже различались светлоты. При мощностях обоих излучений, подобранных должным образом, глаз видит мелькающие поля одинаковой светлоты.
Определив значения относительной спектральной световой эффективности, можно построить кривую ?? = f(?), необходимую для перехода от лучистого потока к световому и обратно (рис. 2.5). Значения ??, стандартизованы (ГОСТ 11093—64).
Спектральная чувствительность палочкового и колбоч-кового аппарата различна. Из рис. 2.5 видно, что кривая относительной спектральной световой эффективности дневного зрения смещена относительно кривой сумеречного зрения.
2.2.2. Адаптация
К свойствам, характеризующим воспроизводящую систему, мы отнесли степень постоянства ее чувствительности.
Чувствительность (световая, спектральная, контрастная и др.) зрительной системы не постоянна, а зависит от ряда факторов, из которых существенное значение имеет уровень освещенности.
Как видно из табл. 2.1, глаз сохраняет чувствительность при разных условиях освещения. Освещенность пейзажа, например при летнем солнце, в 4000 раз больше освещенности, создаваемой в комнате 100-ваттной лампой. Между тем глаз в обоих случаях хорошо различает детали объектов. Человек вполне удовлетворительно видит при освещенности в 1 лк, создаваемой ночью уличными фонарями, т. е. при освещенности в 100 000 раз меньшей, чем при ярком солнце. Глаз дает множество сведений и в полнолуние, когда освещенность почти в 1 000 000 раз ниже, чем при солнце. В безлунную ночь при свете звезд зрение позволяет человеку ориентироваться, хотя освещенность при этом в сотни мил-
лионов раз ниже, чем при открытом солнце.
Ширина диапазона мощностей видимых излучений, могу
щих возбуждать световое ощущение, объясняэться способ-
2.1. Освещенность в различных условиях освещения
|
Условия освещения |
Среднее значение освещенности, лк |
|
Земная поверхность ночью в полнолуние То же в сумерках в летний день в тени в летний день на солнце в летний день при сплошной облачности Пол комнаты под лампой накаливания мощностью 100 Вт, висящей на высоте 3 м Тротуар под уличным фонарем Стол, освещаемый настольной лампой мощностью 100 Вт |
0,2 1—500 6000 — 15 000 50000—120000 5000—25 000 20—30 1 — 6 80—200 |
Если, например, облучить сетчатку насыщенным красным, то белое поле в течение времени адаптации видится зеленым.
Изменение чувствительности глаза во времени, или, как говорят, кинетика яркостной адаптации, показано на рис. 2.6. Процесс увеличения чувствительности при переходе от большой яркости к малой называется темновой адаптацией (рис. 2.6, а), если глаз приспосабливается к большой яркости, — световой адаптацией. Из рисунка видно, что изменение чувствительности глаза продолжается довольно длительное время. Для темновой адаптации это 40—60 мин. Скорость световой адаптации зависит от яркости, на которую адаптируется глаз. Чувствительность падает тем быстрее, чем выше эта яркость. К большой яркости наблюдатель привыкает за 4—8 мин.
В основе яркостной адаптации лежат разные механизмы. Один из них называется зрачковым рефлексом. О нем уже упоминалось в разделе 2.1. При уменьшении освещенности диаметр зрачка увеличивается от 2 мм на ярком свету до 10 мм в полутьме. Световой поток, поступающий в глаз, возрастает при этом в 25 раз, т. е. пропорционально
площади зрачка. Соответственно этому увеличивается и чувствительность.
Более мощный механизм адаптации заключается в двойственности световоспринимающей системы глаза. Палочки позволяют отличить белую поверхность от черной при освещенности 10-6 лк, если глаз адаптирован к такой низкой освещенности. Чувствительность же колбочек гораздо ниже.
По мере роста освещенности палочковый механизм постепенно выключается. При освещенностях выше 10-2 лк зрение становится чисто колбочковым.


Рис. 2.6. Кинетика яркостной адаптации:
а — темновая адаптация (1 — адаптации предшествовала большая яркость; 2 — малая предадаптационная яркость); б — световая адаптация; цифры над кривыми указывают предадаптационные яркости в кд • м-2
Механизм цветовой адаптации заключается в уменьшении концентрации зрительного пигмента в тех колбочках, которые особенно интенсивно работают при предадаптационном освещении.
Так, в приведенном выше примере цветовой адаптации происходит уменьшение концентрации пигмента в красночувствительных рецепторах. Вследствие этого при рассматривании белого поля будут работать главным образом зеленочувствительные и синечувствительные колбочки и глаз получит ощущение зелено-голубого.
Установившийся и неустановившийся зрительный процесс. Чувствительность глаза, как это видно из рис. 2.6, изменяется в процессе яркостной адаптации. В процессе цветовой адаптации изменяется цветовая чувствительность глаза. Зрительный процесс в условиях изменяющейся чувствительности называется неустановившимся. В момент окончания адаптации чувствительность глаза становится постоянной, зрительный процесс при этом носит название установившегося.
Все цветовые измерения и исследования проводятся в условиях установившегося зрения (если, конечно, не изучается сам адаптационный процесс). Поэтому весь дальнейший материал относится к случаям, когда адаптация закончена.
2.2.3. Зрительная инерция
Зрительное ощущение вызывается световым сигналом не мгновенно, а через некоторое время после его начала, и, наоборот, световое ощущение продолжается и после сигнала. Свойство зрительной системы сохранять состояние покоя или работы в течение некоторого времени после начала или
прекращения светового сигнала называется з р-и-тельной инерцией. На рис. 2.7 показано соотношение между постоянным по яркости световым сигналом (тонкая линия) и вызываемым им световым ощущением, выражаемым светлотой (толстая линия). Как видно из рисунка, ощущение светлоты возникает через некоторый момент ?t после начала действия света. На такое же время смещено
и начало спада светлоты. Кривая изменения светлоты во времени отчетливо делится на две части — одна описывает возрастание светлоты, а другая — ее спад в зависимости от времени. Обе части кривой по форме близки к экспоненте. Следствием зрительной инерции является слитное восприятие серии световых сигналов при их достаточно большой частоте.
Если через достаточно малое время вслед за первым световым сигналом последует второй (пунктирные линии на рис. 2.7), то вызванное им ощущение в той или иной мере сольется с первым. Чем меньше промежуток времени, разделяющий сигналы, тем менее прерывисто световое ощущение. При некоторой частоте, когда падение светлоты ?w не превосходит порогового значения, серия сигналов воспринимается как один непрерывный. Частота, обеспечивающая непрерывное зрительное восприятие прерывистых сигналов, называется критической частотой мелькания.
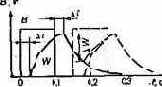
Рис. 2.7. К понятию «зрительная инерция»; штриХпунктирная линия показывает сумму светлот, возникающих вследствие действия двух последовательных световых сигналов, разделенных малым промежутком времени
Вследствие инерции зрения зрительный образ сохраняется в сознании некоторое время после того, как прекратилось действие света. Этот образ называется последовательным. Его возникновение объясняется тем, что продукты фотораспада зрительного пигмента восстанавливаются не сразу после прекращения освещения и отрицательные ионы продолжают давать импульсы тока, приводящие к возникновению зрительного ощущения.
На зрительной инерции основана кинематография. Когда частота кадров равна или превышает критическую частоту мелькания, последовательные образы каждого из кадров сливаются в единое впечатление.
Вследствие зрительной инерции после прекращения ос вещения сохраняется не только ощущение светлоты, но и цветности (см. об этом в разделе 4.1).
2.3. ОСНОВЫ ТЕОРИИ ЦВЕТОВОГО ЗРЕНИЯ
2.3.1. Общие сведения
Характер цветового ощущения находится в связи с распределением чувствительности рецепторов глаза по спектру. Их спектральные свойства изучались многими исследователями, начиная с Максвелла и Кенига, работы которых были выполнены в прошлом веке, и Айвса (1912 г.).
Данные, близкие к современным, были получены Н.Т. и В. Ф. Федоровыми. В настоящее время наиболее надежными считаются значения спектральной чувствительности, определенные Е.
Н. Юстовой (1950 г.) и приведенные на рис. 2.8,

Рис. 2.8. Кривые основных возбуждений (по Е. Н. Юстовой)
а также в таблице 2.2. К ним близки данные Том-сона и Райта (1953 г.).
Графики, изображенные на рисунке, не строго выражают спектральную чувствительность цветочувствительных рецепторов. Они получены для углов зрения, равных 2°. Если принять во внимание малый размер (1,5—2°) участка сетчатки, свободного от палочек, и движение глаз при фиксации взора (см. рис. 2.3), то станет ясным, что при угле зре-
2.2. Ординаты кривых основных возбуждений
|
?, нм |
RК |
R3 |
RС |
?, нм |
Rk |
R3 |
RС |
|
380 |
0,0000 |
0,0000 |
0,0065 |
610 |
0,6864 |
0,2258 |
0,0003 |
|
390 |
0,0001 |
0,0001 |
0,0201 |
620 |
0,5485 |
0,1273 |
0,0002 |
|
400 |
0,0002 |
0,0003 |
0,0679 |
630 |
0,3960 |
0,0663 |
0,0000 |
|
410 |
0,0004 |
0,0011 |
0,2074 |
640 |
0,2686 |
0,0330 |
|
|
420 |
0,0013 |
0,0042 |
0,6456 |
650 |
0,1672 |
0,0157 |
|
|
430 |
0,0036 |
0,0154 |
1 ,3856 |
660 |
0,0963 |
0,0075 |
|
|
440 |
0,0081 |
0,0351 |
1,7471 |
670 |
0,0508 |
0,0035 |
|
|
450 |
0,0142 |
0,0636 |
1,7721 |
680 |
0,0271 |
0,0017 |
|
|
460 |
0,0242 |
0,1049 |
1,6692 |
690 |
0,0131 |
0,0007 |
|
|
470 |
0,0441 |
0,1554 |
1,2876 |
700 |
0,0065 |
0,0004 |
|
|
480 |
0,0817 |
0,2224 |
0,8130 |
710 |
0,0033 |
0 ,0002 |
|
|
490 |
0,1381 |
0,3133 |
0,4652 |
720 |
0,0017 |
0,000* |
|
|
500 |
0,2305 |
0,4647 |
0,2720 |
730 |
0,0008 |
0,0000 |
|
|
510 |
0,3768 |
0,6980 |
0,1582 |
740 |
0,0004 |
|
|
|
520 |
0,5566 |
0,9485 |
0,0782 |
750 |
0,0002 |
|
|
|
530 |
0,7057 |
1,1058 |
0,0422 |
760 |
0,0000 |
|
|
|
540 |
0,8152 |
1,1719 |
0,0203 |
|
|
|
|
|
550 |
0,8903 |
1,1611 |
0,0087 |
|
|
|
|
|
560 |
0,9392 |
1,0865 |
0,0039 |
|
|
|
|
|
570 |
0,9569 |
0,9503 |
0,0021 |
|
|
|
|
|
580 |
0,9408 |
0,7672 |
0,0017 |
|
|
|
|
|
590 |
0,8875 |
0,5621 |
0,0011 |
|
|
|
|
|
600 |
0,8020 |
0,3734 |
0,0008 |
|
|
|
|
ния, равном 2°, могут реагировать не только колбочки, но и палочки. Поскольку нет гарантии, что графики выражают реакции только колбочек, они называются кривыми основных возбуждений (другое название — физиологические кривые сложения — станет ясным из дальнейшего).
Из рисунка видно, что излучения начала видимой части спектра до 430 нм действуют только на синечувствительные колбочки (чувствительности остальных рецепторов в этой области спектра пренебрежимо малы). Их реакция приводит к возникновению ощущения фиолетового цвета.
Различие цветов спектра в пределах 380—430 нм связано только с уровнем реакции рецепторов. Это следует из того, что в указанном диапазоне возбуждаются лишь сине-чувствительные колбочки, но их чувствительность (а следовательно, интенсивность ощущения) возрастает с увеличением длины волны. При продвижении в сторону длинных волн в этом интервале цвета поэтому светлеют. После 430— —440 нм на излучение реагируют и зеленочувствительные рецепторы, поэтому цвет, в зависимости от вклада их реакций в суммарную, постепенно переходит в синий. Затем, приблизительно с 450 нм, наряду с сине- и зеленочувствительными рецепторами начинают работать красночувстви-тельные: синий переходит в голубовато-синий, а затем в голубой. При дальнейшем изменении длины волны цветовое ощущение изменяется по тому же принципу, и относительные значения каждой из реакций на монохроматическое излучение видны из рис. 2.8 и табл. 2.2.
При объяснении возникновения того или иного цветового ощущения приходится сравнивать реакции, даваемые рецепторами разных типов. Например, из рис. 2.8 (или табл. 2.2) видно, что монохроматическое излучение ?, = 650 нм вызывает реакции, находящиеся в отношении R3: RK
= 1:10, и, следовательно, имеет почти чисто-красный цветовой тон. Но сравнение реакций возможно только в том случае, если есть их общая мера. Сложность ее нахождения состоит в том, что реакции, даваемые рецепторами разных типов, качественно различны.
При некоторых соотношениях значений Rk, R3 и Rc
возникает ощущение белого цвета. В этом случае реакции рецепторов всех типов условились считать равными. Они, при определенном их уровне, принимаются за единичные.
Этот принцип используется не только в теории цветового зрения, но и в колориметрии, а также в теории цветовоспроизведения, где количества красок считаются равными, если их наложение дает ахроматический цвет.
Поскольку спектральная чувствительность каждой группы рецепторов выражается одной из кривых основных возбуждений, то можно предположить, что непосредственное сложение кривых дает кривую относительной спектральной эффективности. Однако в результате такого сложения получается несимметричная кривая, не совпадающая с изображенной на рис. 2.5, а кривой. Это объясняется тем, что вклад каждой группы рецепторов в ощущения цветности и светлоты неодинаков. Например, синечувствительные рецепторы очень чувствительны к изменению длины волны, но имеют малую световую чувствительность. Поэтому, для того чтобы перейти от кривых основных возбуждений к кривой относительной спектральной эффективности, нужно ординаты кривых умножить на яркостные коэффициенты. По Е. Н. Юстовой, для Rc яркостный коэффициент равен 0,003, для R3 — 0,65, для RK — 1.
2.3.2. Субъективные характеристики цвета
Характер цветового ощущения зависит как от суммарной реакции цветочувствительных рецепторов, так и от соотношения реакций каждого из трех типов рецепторов. Суммарная реакция определяет светлоту, а соотношение ее долей — цветность.
Хроматические и ахроматические цвета. Когда излучение раздражает все рецепторы одинаково (единица интенсивности раздражения — «доля участия в белом»), его цвет воспринимается как белый, серый или как черный. Белый, серый и черный цвета называются ах роматическими. Эти цвета не различаются качественно. Разница в зрительных ощущениях при действии на глаз ахроматических излучений зависит только от уровня раздражения рецепторов.
Поэтому ахроматические цвета могут быть заданы, одной психологической величиной — светлотой.
Если рецепторы разных типов раздражены неодинаково, возникает ощущение хроматического цвета. Для его описания нужны уже две величины — светлота и цветность. Качественная характеристика зрительного ощущения, определяемая как цветность, двумерна: складывается из насыщенности и цветового тона.
Насыщенность. В тех случаях, когда все рецепторы раздражены почти одинаково, цвет близок к ахроматическому: качество цвета едва выражено. Это, в частности, белый с синим оттенком,- синевато-серый и т. д. Чем больше перевес в раздражении рецепторов одного или двух типов, тем сильнее ощущается качество цвета, его хроматичность. Когда, например, возбуждены только красночувствительные рецепторы, мы видим чисто-красный цвет, весьма далекий от ахроматического.
Степень отличия хроматического цвета от ахроматического называется насыщенностью.
Цветовой тон. Светлота и насыщенность — характеристики, недостаточные для полного определения цвета. Когда говорят «насыщенный красный» или «малонасыщенный зеленый», то, кроме насыщенности, упоминается цветовой тон цвета. Это то его свойство, которое подразумевают в обыденной жизни, когда называют цвет предмета. Несмотря на очевидность понятия, общепризнанного определения термина «цветовой тон» нет. Одно из них дается в такой форме: цветовой тон — это характеристика цвета, определяющая его сходство с известным цветом (неба, зелени, песка и т. д.) и выражаемая словами: «синий, зеленый, желтый и т. д.».
Цветовой тон определяется рецепторами, дающими наибольшую реакцию. Если цветовое ощущение формируется в результате одинакового раздражения рецепторов двух типов при меньшем вкладе третьего, то возникает цвет промежуточного тона. Так, голубой цвет ощущается при одинаковых реакциях зеленочувствительных и синечувствитель-ных колбочек.
Реакция рецепторов, получивших наименьшее раздражение, определяет насыщенность.
Ощущение желтого возникает при равных реакциях красночувствительных и зеленочувствительных колбочек. .Если усиливать возбуждение красночувствительных, цветовой тон смещается в сторону оранжевого. Если вызывать раздражение и синечувствительных, насыщенность упадет.
В главе 3 будет показано, что цветовой тон, насыщенность и светлота данного цвета зависят не только от спектрального состава излучения, но и от условий наблюдения, состояния наблюдателя, цвета фона и т. д. Поэтому рассмотренные здесь характеристики называются субъективными или психологическими.
2.3.3. Действие сложных излучений на рецепторы сетчатки
При действия на глаз смеси излучений реакции рецепторов на каждый из ее компонентов складываются. О цвете смеси можно судить по кривым спектральной чувствительности рецепторов.
Пример 1. Пусть длины волн одинаковых по мощности, например одноваттных, излучений равны:?с — 400 нм, ?3 = 540 нм, ?к = 660 нм. Требуется охарактеризовать цвет смеси.
В этом случае (рис. 2.9) первое излучение (показано сплошными линиями) вызывает реакцию Rc = 0,07, второе — две реакции: R3= 1,17 и RK
= 0,81, а третье R,. = 0,03. Суммарные реакции равны: Rc = 0,07; R3 = 1, 17 и R = = 0,81+0,03 = 0,84. Следовательно, смесь имеет желто-зеленый насыщенный цвет (раздражение синечувствительных рецепторов невелико).
С увеличением мощности раздражителя реакции рецепторов возрастают. Выбирая мощности и длины волн излучений, можно получить самые разнообразные сочетания реакций и, следовательно, ощущение любого цвета.
Пользуясь кривыми основных возбуждений, можно объяснить явление метамерности (метамеризма) цвета тем, что разные сочетания раздражений могут вызвать одинаковые соотношения полных реакций.
Пример 2. Одноваттное монохроматическое излучение ? = 490 нм вызывает реакции, находящиеся в соотношении:

Рис. 2.9. Примеры действия смесей монохроматических излучений на цветочувствительные рецепторы

Рис. 2.10. Схема формирования цветового ощущения: а — кривые основных возбуждений; б—:кривые спектрального распределения мощностей излучения; в — кривые реакций рецепторов
